神経回路の多様性はどこで生まれるのか—三者間シナプスの分子地図—
九州大学高等研究院 生体防御医学研究所 脳機能分子システム分野
発行日:2025年12月30日Published: December 30, 2025
© 2025 日本神経化学会© 2025 The Japanese Society for Neurochemistry
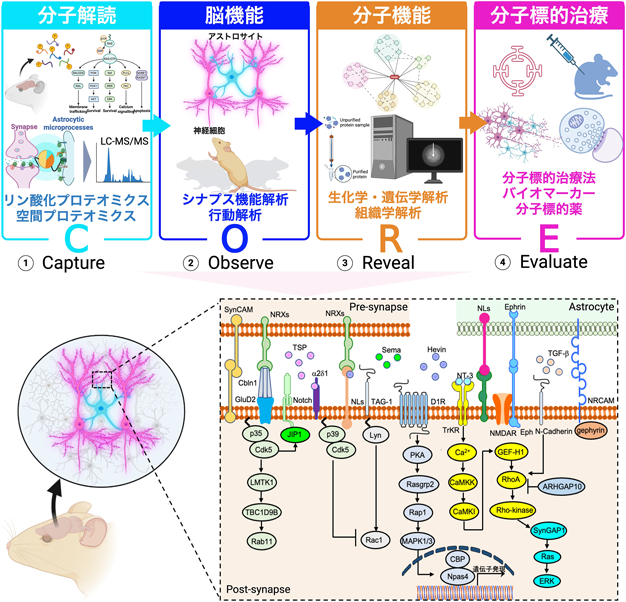
ギリシャ語で「接続」を意味するシナプスは神経細胞間の情報交換を担う主要接触部位であり、生涯にわたる形成・再編・除去の循環によって記憶・学習・情動などの高次機能を支える動的構造である。近年、アストロサイトが三者間シナプス(トリパーティト・シナプス)を構成し、発達期の配線から成体脳の可塑性、興奮/抑制バランス、代謝供給、さらにはシナプス除去に至るまで、近接するシナプスの形成と機能を積極的に統御することが明らかとなった。各シナプスは数千種のタンパク質から成り、その時空間的発現とダイナミクスが小胞放出、受容体リモデリング、接着複合体の再編、シナプス強度の調整を司る。一般意味論を提唱したコージブスキーの「地図は領土ではない」に倣えば、同一の遺伝子セットを共有する細胞型であっても、発達段階・回路文脈・行動状態・疾患病態に応じて、実際に機能するタンパク質の種類・量・役割は大きく変動する。すなわち、個々のシナプスは分子構成と機能の両面で特異化した単位であると捉えられつつあり、神経ネットワークの実在に即して理解するためには、「分子がいつ・どこで・どのように働くか」を同定できる空間解像度と、その機能連関を検証するアプローチを同時に満たすことが不可欠である。さらに、神経変性疾患や精神疾患の初期段階では、三者間シナプスにおける接着・代謝・放出調律の破綻など、シナプス領域のミクロな時空間異常が早期から顕在化する可能性が示唆されている。ゆえに、病態機序の解明と分子標的の創出に向け、シナプス分子多様性の写像とその動態・機能の実証が必須である。筆者らの研究室は、CORE(Capture–Observe–Reveal–Evaluate)を研究の基本コンセプトとして、三者間シナプスの分子多様性を体系的に解析してきた(図1)。Captureでは、近位依存性ビオチン標識(BioID)やリン酸化・空間プロテオミクスを用いて細胞種・回路・時間に応答する分子群を生体内で捕捉し、LC–MS/MSにより網羅的に同定する。Observeでは、アストロサイトを含む回路単位のシナプス機能解析と行動解析を統合して表現型を捉える。Revealでは、生化学・遺伝学・組織学を横断する検証により、候補分子の因果関係と生理機能を明らかにする。Evaluateでは、得られた分子学的知見をバイオマーカー探索や分子標的治療の評価へと橋渡しし、基礎から臨床応用へ展開している。本稿では、これまでの筆者らのシナプス分子地図をめぐる旅の歩みから、神経回路の形成・維持・可塑性を支える包括的分子メカニズムを概説する。
神経回路の基本単位である軸索・樹状突起・スパインは、高度に発達した細胞内小胞輸送に支えられて形成・維持・可塑化される動的構造である1)。なかでもリサイクリングエンドソームは、受容体や接着分子、シナプス小胞関連タンパク質を、必要なときに必要な場所へ届ける細胞内物流輸送の中核ハブである。都市の物流にたとえるなら、宛先情報(ソーティングシグナル)に従い、配達員(輸送小胞/モーター蛋白)が品物(タンパク質・脂質)を運び、交通管制(Rabファミリー)がそれらの流れを調整している。しかし、リサイクリングエンドソームが品物を「いつ・どこへ・どれだけ」運ぶかをいかに決めているのか、その時空間制御の全体像は長らく不明であった。
筆者らは、この制御の上流に位置する因子として機能未知のリン酸化酵素LMTK1(AATYK1)に着目した2, 3)。LMTK1はCDK5依存的リン酸化により活性が変化し、Rab11陽性リサイクリングエンドソームの移動特性(頻度・速度・走行距離)を抑制的に調整することを明らかにした(図1)4)。この細胞内の物流システムにより、過剰な資材(とくに膜脂質)が局所に供給されるのを防ぎ、突起やスパインの資材配分を最適化することで、神経細胞は他の細胞にはない情報伝達に特化した特徴的かつ美麗な形態を獲得する5, 6)。実際に、筆者らは軸索形成・樹状突起形成・スパイン形成の各段階において、LMTK1がRab11小胞の順行輸送を抑制して過伸長を防ぐブレーキとして機能することを示した4–6)。LMTK1機能が低下すると、リサイクリングエンドソームの順行輸送が過剰化し、軸索の異常伸長や分岐が増加することで神経回路の配線エラーのリスクが高まる。これらの結果は、LMTK1がリサイクリングエンドソームを介した脂質・膜資材の供給を時空間的に最適化し、発生期のダイナミックな形態形成を統御していることを示すものである。分子機構としては、LMTK1がRab11の不活性化因子TBC1D9Bを介してRab11活性を抑制することを見出した。すなわち、CDK5–LMTK1–TBC1D9B–Rab11という一つの細胞内シグナル経路が、神経細胞における物流(小胞輸送)のいわば“管制塔”を担うことを初めて明らかにしたのである(図1)1, 5, 6)。この制御系は行動レベルにも投影される。LMTK1遺伝子欠損マウスでは多動性・衝動性の増加が認められ、細胞内小胞輸送の恒常性破綻が精神行動異常へ波及しうることが示された1, 7)。さらに神経変性の発症機構にも関与し、LMTK1がβセクレターゼBACE1のリサイクリングエンドソーム局在を制御し、キナーゼ活性の喪失がこの制御を破綻させることを明らかにした8)。BACE1はアルツハイマー病のアミロイドβ産生・凝集に関与するため、LMTK1機能不全はアミロイドβの形成効率やタイミングを変えうる上流因子となる可能性が高い。以上の結果から、CDK5–LMTK1–TBC1D9B–Rab11経路は、リサイクリングエンドソームの時空間制御を介して神経細胞の形態と機能を協調的に最適化し、その破綻が発達障害に起因する行動異常および神経変性へ連なる分子基盤であることが示された。
現存するすべての形は「材料を足す(伸長)」だけでなく「余分を引く(退縮)」という二つの力学がつくる平衡状態として定まる9, 10)。これは、コンピューターやAIの父として知られるアラン・チューリングが提案した反応拡散モデルにも通じ、局所の活性化と長距離に及ぶ拡散的抑制の拮抗によって秩序ある形態パターンが生じるという原理である。神経細胞の極性形成も同様であり、成長を促す局所シグナルと全細胞内に波及する抑制シグナルの釣り合いが、どの突起を伸ばし、どの突起を退かせるかを選び分け、最終的に一本の軸索を確立させる。具体的には、生まれたばかりの神経細胞は複数の未熟突起をもつが、やがて一本だけが軸索としての機能を獲得し、残りは樹状突起へ分化する。この軸索の一本化に失敗すれば、入力と出力の向きが曖昧となり、発達期の配線エラーに直結する。では、どの突起を伸ばし、どの突起を退かせるのか。その運命決定の基本原理はいかなるものなのだろうか。
筆者らは、先行突起(将来の軸索)が抑制性シグナルを全細胞内へ遠隔送信し、他突起の伸長を選択的に止めるというモデルを提案した9, 10)。合奏に喩えれば、主旋律を際立たせるために指揮者が伴奏を抑える状況である。高時間分解能Ca2+イメージングにより、成長円錐が神経栄養因子(NT-3やBDNF)に応答して大振幅のCa2+波を発し、それが細胞体へ伝搬することを発見した。この波は細胞体でRhoAからRho-kinase(ROCK)へ至る経路を活性化し、第二の軸索形成を抑えるスイッチとして機能する(図1)11)。しかも抑制の大きさは一様ではなく、突起ごとに強さが異なるという空間選択性を示した。筆者らは独自の光分子操作技術(LOVTRAP)を用いてシグナルの発生位置とタイミングを精密に制御し、数理モデル解析と統合して抑制効果の距離依存性を定量化した11)。その結果、細胞体で立ち上がった収縮シグナルの効き目は突起の長さに応じて変化し、短い未熟突起ほど強く抑制されることが明らかとなった。すなわち、先行突起は細胞体というハブを介して距離依存的な抑制を全突起へ伝達し、近傍で軸索化しようとする複数の突起を選択的に退縮させている。
では、この空間的な遠隔抑制シグナルを実装する分子メカニズムは何か。筆者らはリン酸化プロテオミクス(KISS法)を用いて、長距離Ca2+伝搬の下流で作動するリン酸化シグナルネットワークを網羅的に解析した11, 12)。その結果、カルシウムカルモジュリン依存性タンパク質キナーゼCaMKIがRhoAの活性化因子であるGEF-H1をリン酸化して機能を亢進させ、RhoAからRho-kinaseへ至る抑制性機能を駆動していた。これによりアクチン骨格の収縮が誘導され、競合突起の退縮が起こる。以上より、遠隔抑制型シグナル伝達(長距離Ca2+伝搬→CaMKI→GEF-H1→RhoA→Rho-kinase)が、第二の軸索形成を抑え、回路の一方向性の情報伝達を生み出す神経細胞固有の形態決定機構であることが示唆された(図1)11)。
脳では、シナプスとアストロサイトがつくる「三者間シナプス」において、代謝供給、受容体配置、可塑性の閾値が協働して定まる13)。にもかかわらず、どの分子が・どの場所で・どのように働くのかを、生体内で回路文脈を保ったまま特定することは容易ではない14, 15)。免疫染色やバルクのプロテオームでは、細胞間で生じる数十nm規模の相互作用を選択的に切り出しにくく、単一の手法で特異性と網羅性を同時に満たすのは測定学的ジレンマであった。筆者らは、この壁を越えるために、アストロサイトとニューロンが実際に触れ合う接着部位そのものを「刻印」して分子実態を解読する空間プロテオーム解析Split-TurboIDとTurboID-surfaceを開発した13–16)。Split-TurboIDでは、近接標識酵素TurboIDを二分割し、一方をアストロサイトの細胞外面、他方をニューロンの細胞外面に提示する。二つの細胞が接触したときにのみ断片が再会合して活性化し、半径数~十数nmの範囲にある表層タンパク質をビオチン標識する。すなわち、三者間シナプス部位に対して極めて選択的に刻印する仕組みである。一方のTurboID-surfaceは、GPIアンカーや膜貫通配列を用いてTurboIDをアストロサイト表層に広く提示し、受容体・接着分子・輸送体などの膜タンパク質を包括的に標識するラベルとなる。両者を重ね合わせることで、特定の細胞種間に実在する分子ネットワークを高信頼に抽出できる。
これらの空間プロテオーム解析技術の開発には、度重なる試行と改良の積み重ねによって確立された。酵素選定では当初BioID2を用いたが、生体組織内での活性不足が壁となった。しかし、米国BRAIN Initiative会議での発表を契機に手法の情報共有を進め、本アプローチは各研究グループへ波及した。Split-TurboIDの最適化では、非接触時の自発的再会合を抑えつつ、接触時には確実に再会合させるという相反条件の均衡点を探り、リンカー長を調整した。短すぎれば感度が落ち、長すぎれば離れた分子を誤標識する。最終的に、膜直下に最小限の柔軟性を残す設計とし、酵素由来の人工接着やシナプス密度の乱れを回避した。TurboID-surfaceでも、初期設計の一部で樹状突起近傍における内在化が目立ち表層解像度が低下したため、GPIアンカーへの切替とバックグラウンド補正の強化で改善した。分子同定は二段階のプロテームの照合で一貫して絞り込んだ。第一に、Split-TurboIDで得た候補がTurboID-surfaceでも検出されるかを確認し、双方で同定された分子を三者間シナプス構成分子とした。第二に、TurboID-surfaceのみで検出されSplit-TurboIDで検出されない分子は、三者間シナプス以外のアストロサイト細胞膜表層分子と定義した。標識タンパク質をストレプトアビジンで精製し、LC–MS/MSにより同定・定量した結果、計3,171種のタンパク質由来ペプチドを取得した。厳密な閾値(1.5倍濃縮)で候補を抽出し、両法に共通する118種を高信頼の三者間シナプス・プロテオームとして定義した16)。分子プロファイルでは既知の細胞間隙分子やイオンチャネルに加え、細胞接着分子の豊富さが際立った。中でも、筆者らは新たな三者間シナプス分子NRCAMに着目した。NRCAMは皮質アストロサイトに発現し、三者間シナプス接触部に局在すること、さらに免疫グロブリン様ドメインがアストロサイト形態の維持に必須であること(欠失変異ではレスキュー不能)を示した。超解像顕微鏡STEDと電気生理解析の結果、アストロサイトにおけるNRCAM欠損は抑制性シナプスとの接触を選択的に障害し、mIPSCの頻度・振幅を低下させる一方、興奮性入力は相対的に保たれた。分子機序としては、トランスのNRCAM–NRCAM結合がニューロン側gephyrin複合体と相互作用し、抑制性シナプスの形成を支えることが分かった(図1)16)。以上より、Split-TurboIDとTurboID-surfaceを統合する空間プロテオミクスは、三者間シナプス間隙において「どの分子が・どの場所で・どのように働くか」を、生体内でかつ回路文脈のまま解像するための実証的基盤を与えた13–16)。さらに、本研究では分泌因子中心の従来像に、接着コードという新たな枠組みを組み込み、とりわけ抑制性シナプスの選択的編成を支える全く新しい作動原理を提示した。
本稿では、単一シグナル経路から網羅的シグナル解析、さらに空間プロテオーム解析へと展開してきた筆者らの研究、いわば一本の木から森全体へ視座を広げる歩みとして概観した。これらの大規模な分子解析から見えてくるのは、タンパク質はけして単独で働くのではなく、時間と空間に配列された分子ネットワークとして機能している点である。実際、ヒトには約十万種のタンパク質が存在し、それらが適所への配置と複合体形成を通じて多彩な生理機能を担う。ゆえに、心や記憶の原理を理解するためには、タンパク質の全体的な空間構成と機能、ならびに分子ネットワークの文脈を解読することが不可欠である。冒頭でも述べたコージブスキーの「地図は領土ではない」に倣えば、遺伝子発現やバルク量という地図から実在の回路機能という領土を読み解くには、時間、空間、相互作用の三座標を備えた新たなプロテオーム情報が必要となる。筆者らが開発してきた空間プロテオーム解析は、この不足を補う分子基盤となりうる。一方で、未解決課題も明確である。第一に、空間プロテオームは脳内分子ネットワークの“どこに在るか”を示すが、“どれだけ効いているか”を定量するには、遺伝学的介入、光操作、生理計測、機械学習を統合する複合的解析が必須である。第二に、シナプスは時間とともに変化するため、発達・学習・睡眠・ストレス・病態進行を横断する縦断解析によって、分子ネットワークの軌跡と可逆性/不可逆性を見極める必要がある。第三に、三者間シナプスにとどまらず、ミクログリアやオリゴデンロサイトを含むグリア多様性や微小血管との細胞接着部位など、未踏の接着コードがなお多く残されている。これらに迫るには、時相選択的近接標識、単一シナプス解像度のプロテオミクス、シングルセル・トランスクリプトーム/メタボローム/脂質オミクスを統合するマルチオミクス解析が不可欠である。とりわけ重要なのは、得られた分子ネットワークを再現可能な形式で公開し、分野横断で再利用できる分子地図として整備することである。忘れてはならないのは、分子地図の精緻化それ自体が目的ではないという点である。目指すべきは、回路が生み出す行動と認知という実在の理解を深め、その知見を診断や介入へ橋渡しすることである。とくに、発達障害や精神・神経疾患を含む多くの病態では、回路特異的標的分子に立脚した創薬が現実味を帯びる。個々の神経回路レベルでの高精度分子マッピングは、抗体医薬や分子標的薬の開発に直結し、患者ごとの回路特性に即した治療設計を促して、より有効で副作用の少ない個別化医療の実装に繋がるものと期待される。
本研究の遂行にあたり、藤田医科大学の貝淵弘三教授、東京都立大学の久永眞市教授、Duke UniversityのScott Soderling教授、慶應義塾大学の柚﨑通介教授より、多大なるご指導を賜りました。ここに厚く御礼申し上げます。あわせて、東北大学大学院の福田光則教授、名古屋大学の本田直樹教授、明海大学の友村美根子教授には多大なご助力を賜りました。心より感謝申し上げます。また、九州大学・脳機能分子システム分野ならびに分子神経免疫学分野の研究室メンバーの皆様からは、有益なご助言と技術的ご支援を頂戴いたしました。ここに深く感謝申し上げます。最後に、本稿執筆の機会をお与えくださいました日本神経化学会理事長の小泉修一先生、優秀賞・奨励賞選考委員会委員長の津田誠先生、並びに委員会の先生方に、謹んで御礼申し上げます。
1) Hisanaga SI, Wei R, Huo A, Tomomura M. LMTK1, a novel modulator of endosomal trafficking in neurons. Front Mol Neurosci, 13, 112 (2020).
2) Tsutsumi K, Takano T, Endo R, Fukuda M, Ohshima T, Tomomura M, Hisanaga S. Phosphorylation of AATYK1 by Cdk5 suppresses its tyrosine phosphorylation. PLoS One, 5(4), e10260 (2010).
3) Takano T, Tsutsumi K, Saito T, Asada A, Tomomura M, Fukuda M, Hisanaga S. AATYK1A phosphorylation by Cdk5 regulates the recycling endosome pathway. Genes Cells, 15(7), 783–797 (2010).
4) Takano T, Urushibara T, Yoshioka N, Saito T, Fukuda M, Tomomura M, Hisanaga S. LMTK1 regulates dendritic formation by regulating movement of Rab11A-positive endosomes. Mol Biol Cell, 25(11), 1755–1768 (2014).
5) Takano T, Tomomura M, Yoshioka N, Tsutsumi K, Terasawa Y, Saito T, Kawano H, Kamiguchi H, Fukuda M, Hisanaga S. LMTK1/AATYK1 is a novel regulator of axonal outgrowth that acts via Rab11 in a Cdk5-dependent manner. J Neurosci, 32(19), 6587–6599 (2012).
6) Nishino H, Saito T, Wei R, Takano T, Tsutsumi K, Taniguchi M, Ando K, Tomomura M, FUKUDA M, Hisanaga SI. The LMTK1-TBC1D9B-Rab11A cascade regulates Dendritic spine formation via endosome trafficking. J Neurosci, 39(48), 9491–9502 (2019).
7) Takahashi M, Sugiyama A, Wei R, Kobayashi S, Fukuda K, Nishino H, Takahashi R, Tsutsumi K, Kita I, Ando K, Manabe T, Kamiguchi H, Tomomura M, Hisanaga SI. Hyperactive and impulsive behaviors of LMTK1 knockout mice. Sci Rep, 10(1), 15461 (2020).
8) Komaki K, Takano T, Sato Y, Asada A, Ikeda S, Yamada K, Wei R, Huo A, Fukuchi A, Saito T, Ando K, Murayama S, Araki W, Kametani F, Hasegawa M, Iwatsubo T, Tomomura M, Fukuda M, Hisanaga SI. Lemur tail kinase 1 (LMTK1) regulates the endosomal localization of β-secretase BACE1. J Biochem, 170(6), 729–738 (2022).
9) Takano T, Funahashi Y, Kaibuchi K. Neuronal polarity: Positive and negative feedback signals. Front Cell Dev Biol, 7, 69 (2019).
10) Takano T, Xu C, Funahashi Y, Namba T, Kaibuchi K. Neuronal polarization. Development, 142(12), 2088–2093 (2015).
11) Takano T, Wu M, Nakamuta S, Naoki H, Ishizawa N, Namba T, Watanabe T, Xu C, Hamaguchi T, Yura Y, Amano M, Hahn KM, Kaibuchi K. Discovery of long-range inhibitory signaling to ensure single axon formation. Nat Commun, 8(1), 33 (2017).
12) Nagai T, Nakamuta S, Kuroda K, Nakauchi S, Nishioka T, Takano T, Zhang X, Tsuboi D, Funahashi Y, Nakano T, Yoshimoto J, Kobayashi K, Uchigashima M, Watanabe M, Miura M, Nishi A, Kobayashi K, Yamada K, Amano M, Kaibuchi K. Phosphoproteomics of the dopamine pathway enables discovery of Rap1 activation as a reward signal in vivo. Neuron, 89(3), 550–565 (2016).
13) Takano T, Soderling SH. Tripartite synaptomics: Cell-surface proximity labeling in vivo. Neurosci Res, 173, 14–21 (2021).
14) Ito Y, Nagamoto S, Takano T. Synaptic proteomics decode novel molecular landscape in the brain. Front Mol Neurosci, 17, 1361956 (2024).
15) Matsubayashi J, Takano T. Proximity labeling uncovers the synaptic proteome under physiological and pathological conditions. Front Cell Neurosci, 19, 1638627 (2025).
16) Takano T, Wallace JT, Baldwin KT, Purkey AM, Uezu A, Courtland JL, Soderblom EJ, Shimogori T, Maness PF, Eroglu C, Soderling SH. Chemico-genetic discovery of astrocytic control of inhibition in vivo. Nature, 588(7837), 296–302 (2020).

This page was created on 2025-12-18T10:26:41.62+09:00
This page was last modified on 2026-01-06T16:27:59.000+09:00
このサイトは(株)国際文献社によって運用されています。