液–液相分離によるシヌクレイノパチー発症機構の解明
熊本大学発生医学研究所ゲノム神経学分野
発行日:2025年12月30日Published: December 30, 2025
© 2025 日本神経化学会© 2025 The Japanese Society for Neurochemistry
αシヌクレイン(α-synuclein: αSyn)は、パーキンソン病やレビー小体型認知症に代表されるシヌクレイノパチー患者脳における特徴的な病理所見である「細胞内凝集体」の主要構成タンパク質である。αSynは、正常な神経細胞ではシナプス小胞と細胞膜の融合に関与するSNAREタンパク質複合体の一部としてシナプス前終末に局在する。一方、シヌクレイノパチー患者脳では何らかの異なる分子と複合体を形成し、細胞内にアミロイド凝集体を形成する。近年、αSynアミロイド凝集体は分子クラウディングの一形態である液−液相分離(liquid-liquid phase separation: LLPS)を介して形成されることが報告されているが、これは主にαSynタンパク質単体を用いたin vitro研究にとどまっており1)、in vivoでの病態発症メカニズムとの関連は未だ不明である。
一般に広く知られている核酸の構造は右巻きの二重らせん構造であり、この基本構造は「B型」と呼ばれている2)。このB型構造以外にも、左巻きらせん構造であるZ型や、三重鎖構造(H型)など、いわゆる「非B型」構造と総称される多様な高次構造が存在することが明らかになっている3)。これらの構造は、塩基配列の特徴や溶液条件などの環境要因に応じて形成されることが知られている。非B型核酸構造の一つであるグアニン四重鎖(G-quadruplex: G4)は、グアニンに富む配列を有する一本鎖核酸上に形成される高次構造である(図1)。本稿では、RNAグアニン四重鎖(RNA G-quadruplex: G4RNA)について概説するとともに、我々が最近明らかにしたG4RNAを介したαSyn相転移制御機構と神経変性メカニズムについて紹介する。
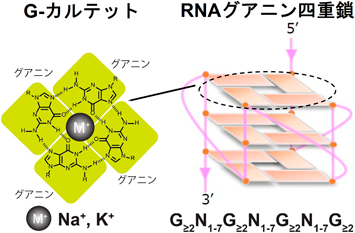
各グアニン塩基は隣接する2つのグアニンと水素結合を形成し、4つのグアニンが「Gカルテット」と呼ばれる平面構造をとる。このGカルテットが積み重なることで、G4構造が構築される。G4構造は、4つのグアニントラクトとそれらを連結する3つのループ領域によって形成され、ループの長さや配列の違いによって多様な構造多様性が生じる。特に、RNA由来のG4(rG4)は、すべてのストランドが同一方向を向く「パラレル型トポロジー」をとることが知られている(文献4より改変)。
G4構造は各グアニン塩基が隣接する2つのグアニン塩基と非ワトソン・クリック型のフーグスティーン塩基対を形成することで、4つのグアニンが正方形状に配列されたGカルテットを構成する(図1)。このGカルテットが複数積み重なることにより、G4構造が形成される。G4は、少なくとも2つの連続したグアニンを含む4つのグアニントラクトと、それらを連結する3つのループ領域から構成される(図1)4)。G4の形成および安定性は、カチオンの種類、温度、ストランドの配向、ループの長さなど、複数の要因によって影響を受ける5)。特に、Gカルテットの中心に一価の陽イオンが結合することで構造が安定化されることが知られており、一般的にはK+>Na+>Li+の順に安定化作用が強いとされている6)。ループの長さや配列により多様なG4構造が形成されるが、G4RNAは、ループ長にかかわらずストランドがすべて同一方向を向くパラレル型トポロジーをとることが特徴である(図1)7, 8)。一方で、これらG4構造のトポロジーに関する知見は、すべてin vitro解析に基づくものであり、その物理化学的特性がin vivoにおける生理機能にどの程度寄与しているのかについては依然として不明な点が多い。
まず我々はαSynが神経細胞内でLLPSを介して凝集するかどうかを検討した。シヌクレイノパチー患者の死後脳ではリン酸化αSyn(pS129)陽性の凝集体が観察されており、pS129はαSyn凝集のマーカーとして広く用いられている。αSynアミロイド線維(preformed fibril: PFF)をマウス神経細胞に処置すると、時間経過に伴って患者脳と同様のpS129陽性凝集体が細胞体に出現することが知られている。一方で、pS129陽性凝集体が形成される前段階におけるαSynの細胞内動態は不明であった。我々は、ヒト由来のαSyn PFFをマウス培養神経細胞に処置し、マウスαSynに特異的な抗体を用いてその動態を観察した。その結果、αSynはまずDCP1a陽性のRNA顆粒(P-body)に集積し、その後にpS129陽性の凝集体を形成することが明らかとなった。P-bodyを含む非膜オルガネラは、多価の分子間相互作用によって形成されるLLPSによって構築されることが知られている。LLPSはタンパク質や核酸が多価の相互作用によって液滴状に会合した状態であることから9)、αSynの物性に対するタンパク質およびRNAの影響についても検討した。マウス神経芽細胞腫から抽出した細胞由来タンパク質またはRNAをin vitroの分子クラウディング条件下で精製αSynと混合したところ、細胞由来タンパク質ではαSyn液滴に顕著な影響は見られなかったが、細胞由来RNAを加えるとαSynはゾル–ゲル相転移を呈した。αSynのRNA結合性についてはこれまで不明であったため、RNA-Bind-n-seq法を用いてランダム配列RNAの中からαSynが結合する配列を同定した10)。その結果、αSynはグアニンが豊富で連続する一次配列を有するRNAによく結合することが示された。このような配列では先述したG4構造の形成が示唆されるため、αSynのRNA結合においてG4RNAが関与するかを検証するためにゲルシフト解析を実施した。代表的なG4RNAであるテロメアRNA(UUAGGG)4、G4構造を形成しない変異型テロメアRNA(UUACCG)4、ミスマッチヘアピン構造(CAG)8、およびポリアデニン(AAA)8を用いて比較した結果、G4RNAのバンドはαSyn濃度に依存して上方にシフトし、他のRNA配列では明確な移動が見られなかった。この結果は、αSynがG4RNAに特異的に結合することを示している。さらに、G4RNAを加えた条件下でαSyn液滴の状態を観察すると、予想通りαSynはゾル–ゲル相転移を示した。これらの結果から、G4RNAはαSynの凝集を誘導するキーファクターであることが示唆された。
次に、細胞ストレス時におけるG4RNAの動態について検討した。マウス培養神経細胞にαSyn PFFを処置したところ、pS129陽性凝集体が形成される以前に、G4RNAの増加および会合が観察され、最終的にはG4RNAがαSynと共に凝集することが明らかとなった。さらに、このようなαSynとG4RNAの共凝集体はパーキンソン病患者の死後脳組織においても確認された。興味深いことに、G4RNAの増加および会合はカルシウムイオノフォアであるイオノマイシン処置によって誘導され、in vitroにおいてもカルシウムイオン(Ca2+)濃度依存的にG4RNAの自己会合による相分離が促進されることが確認された。また、Ca2+存在下では、G4RNAによるαSynのゾル–ゲル相転移が強く誘導された。加えて、αSyn PFF処置後の神経細胞においてはカルシウムホメオスタシスが破綻し、細胞内Ca2+濃度が高く保たれていることが明らかとなった。これらの結果は、Ca2+によるG4RNAの自己会合がαSynのゾル–ゲル相転移の起点となる可能性を示唆している。
続いて、細胞内におけるG4RNA会合がαSynの凝集を誘導するかどうかを検討した。細胞内でG4RNAの自己会合を制御可能な実験系として、光照射によってG4RNA会合を誘導するOptoG4システムを構築した。このシステムでは、G4RNA形成配列にバクテリオファージMS2由来のステムループ構造(MS2配列)を付加したコンストラクトとMS2ステムループに特異的に結合するMS2コートタンパク質(MCP)に、光照射で自己会合を誘導するCry2oligoを融合させたコンストラクトを細胞に発現させ、青色光(480 nm)を照射することで細胞内におけるG4RNAの会合を人為的に制御可能とした。このOptoG4システムをマウス培養神経細胞に導入し、青色光を照射すると、G4RNAの会合に伴いαSynが共凝集し、さらに興奮性神経伝達の障害が引き起こされた。この神経機能障害はG4構造を形成しないRNA配列を用いた場合には観察されなかった。次に、in vivoでの検討を行うため、アデノ随伴ウイルスを用いてマウスの黒質ドパミン神経細胞にOptoG4システムを特異的に発現させて青色光を照射した。その結果、OptoG4発現マウスでは時間経過に伴ってパーキンソン病様の運動機能障害が観察された。作製した脳スライスにおいてドパミン神経細胞を観察すると、in vitroと同様にG4RNAとαSynの共凝集体が確認され、ドパミン神経細胞の変性および脱落が認められた。以上の結果から、G4RNAの自己会合が細胞内におけるαSyn凝集の起点となり、それが神経変性を誘導することが示された11)。
最後に、αSyn凝集に寄与する内在性G4RNAについて、免疫沈降法とRNAシーケンスを組み合わせた解析を行った12)。G4構造認識抗体およびpS129抗体を用いて免疫沈降を行い、濃縮されたRNAをシーケンス解析したところ、その大半がシナプス関連タンパク質をコードするmRNAであることが判明した。特に、CaMKIIやPSD95といったシナプス可塑性に必須なmRNAが含まれており、これらは3′-UTR領域にG4構造を形成することが既に知られている。これらの結果は、細胞ストレスによりシナプス関連分子のmRNA上に形成されたG4構造がLLPSを介して自己会合し、それがαSyn凝集の足場となる可能性を示唆している。さらに、G4RNAとαSynの共凝集によりRNAの隔離が起こり、それに伴って局所翻訳が障害され、結果としてシナプス機能不全が引き起こされることが考えられる(図2)。
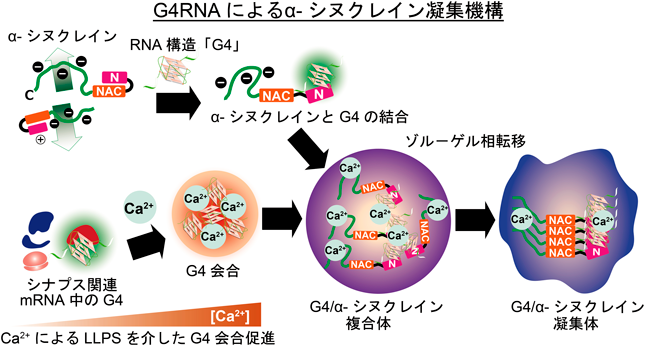
通常、αSynはモノマーとして機能している。細胞ストレスにより細胞内カルシウムイオン濃度が上昇すると、G4RNAが増加・会合し、その結果としてαSynはLLPSを介してゾル–ゲル相転移を起こし、凝集体を形成する。αSyn凝集に関与するG4RNAの多くはシナプス関連分子をコードするmRNAであり、これらのmRNAとαSynが共凝集することでタンパク質翻訳が阻害され、シナプス機能が障害される。最終的に神経変性が引き起こされ、神経変性疾患の発症に至ると考えられる(文献11より改変)。
本稿では、G4RNAがαSynのゾル–ゲル相転移の起点となり、シヌクレイノパチー発症に寄与することを紹介した11)。本稿では詳細を割愛するが、G4RNAがαSynのN末端に結合してゾル–ゲル相転移を誘導すること(図2)、また、G4構造を不安定化させる化合物がパーキンソン病様の神経変性を予防しうるという結果も得ている。さらに、G4RNAはαSynだけでなく、Tauを含む他のプリオン性タンパク質のゾル–ゲル相転移にも寄与しており13, 14)、多くの神経変性疾患の発症にG4構造が関与している可能性が示唆される。今後は、G4RNAによる神経変性機構について、疾患間での「共通性」と「特異性」を明らかにするとともに、G4構造を標的とした新たな治療法および予防法の開発に向けて、精力的に研究を進めていきたい。
本研究は、熊本大学発生医学研究所ゲノム神経学分野・塩田倫史教授のご指導のもとに実施することができました。ここに深く感謝申し上げます。また、多くの共同研究者の先生方ならびに、日本医療研究開発機構をはじめとする各種研究助成により多大なご支援を賜りましたこと、心より御礼申し上げます。さらに、本研究成果をご評価いただき、2025年度 日本神経化学会優秀賞に選出いただきましたこと、関係者の皆様に厚く御礼申し上げます。
1) Ray S, Singh N, Kumar R, Patel K, Pandey S, Datta D, Mahato J, Panigrahi R, Navalkar A, Mehra S, Gadhe L, Chatterjee D, Sawner AS, Maiti S, Bhatia S, Gerez JA, Chowdhury A, Kumar A, Padinhateeri R, Riek R, Krishnamoorthy G, Maji SK. α-Synuclein aggregation nucleates through liquid-liquid phase separation. Nat Chem, 12(8), 705–716 (2020).
2) Watson JD, Crick FH. Molecular structure of nucleic acids; a structure for deoxyribose nucleic acid. Nature, 171(4356), 737–738 (1953).
3) Zhao J, Bacolla A, Wang G, Vasquez KM. Non-B DNA structure-induced genetic instability and evolution. Cell Mol Life Sci, 67(1), 43–62 (2010).
4) Asamitsu S, Takeuchi M, Ikenoshita S, Imai Y, Kashiwagi H, Shioda N. Perspectives for applying G-quadruplex structures in neurobiology and neuropharmacology. Int J Mol Sci, 20(12), 2884 (2019).
5) Kumar N, Sahoo B, Varun KA, Maiti S, Maiti S. Effect of loop length variation on quadruplex-Watson Crick duplex competition. Nucleic Acids Res, 36(13), 4433–4442 (2008).
6) Bhattacharyya D, Mirihana Arachchilage G, Basu S. Metal cations in G-quadruplex folding and stability. Front Chem, 4, 38 (2016).
7) Patel DJ, Phan AT, Kuryavyi V. Human telomere, oncogenic promoter and 5′-UTR G-quadruplexes: diverse higher order DNA and RNA targets for cancer therapeutics. Nucleic Acids Res, 35(22), 7429–7455 (2007).
8) Zhang AY, Bugaut A, Balasubramanian S. A sequence-independent analysis of the loop length dependence of intramolecular RNA G-quadruplex stability and topology. Biochemistry, 50(33), 7251–7258 (2011).
9) Ryan VH, Fawzi NL. Physiological, pathological, and targetable membraneless organelles in neurons. Trends Neurosci, 42(10), 693–708 (2019).
10) Lambert N, Robertson A, Jangi M, McGeary S, Sharp PA, Burge CB. RNA Bind-n-Seq: quantitative assessment of the sequence and structural binding specificity of RNA binding proteins. Mol Cell, 54(5), 887–900 (2014).
11) Matsuo K, Asamitsu S, Maeda K, Suzuki H, Kawakubo K, Komiya G, Kudo K, Sakai Y, Hori K, Ikenoshita S, Usuki S, Funahashi S, Oizumi H, Takeda A, Kawata Y, Mizobata T, Shioda N, Yabuki Y. RNA G-quadruplexes form scaffolds that promote neuropathological α-synuclein aggregation. Cell, 187(24), 6835–6848.e20 (2024).
12) Asamitsu S, Yabuki Y, Matsuo K, Kawasaki M, Hirose Y, Kashiwazaki G, Chandran A, Bando T, Wang DO, Sugiyama H, Shioda N. RNA G-quadruplex organizes stress granule assembly through DNAPTP6 in neurons. Sci Adv, 9(8), eade2035 (2023).
13) Asamitsu S, Yabuki Y, Ikenoshita S, Kawakubo K, Kawasaki M, Usuki S, Nakayama Y, Adachi K, Kugoh H, Ishii K, Matsuura T, Nanba E, Sugiyama H, Fukunaga K, Shioda N. CGG repeat RNA G-quadruplexes interact with FMRpolyG to cause neuronal dysfunction in fragile X-related tremor/ataxia syndrome. Sci Adv, 7(3), eabd9440 (2021).
14) Yabuki Y, Matsuo K, Komiya G, Kudo K, Hori K, Ikenoshita S, Kawata Y, Mizobata T, Shioda N. RNA G-quadruplexes and calcium ions synergistically induce Tau phase transition in vitro. J Biol Chem, 300(12), 107971 (2024).

This page was created on 2025-11-27T16:11:31.317+09:00
This page was last modified on 2026-02-18T13:44:40.000+09:00
このサイトは(株)国際文献社によって運用されています。