リソソーム膜破綻に起因する異常タンパク凝集の伝播とその防御機構に関する研究
大阪大学大学院医学系研究科 神経内科学教室
発行日:2025年12月30日Published: December 30, 2025
© 2025 日本神経化学会© 2025 The Japanese Society for Neurochemistry
近年、高齢化の進行に伴い神経変性疾患の患者数が急増しており社会的に深刻な課題となっている。神経変性疾患は中枢神経系の特定の神経細胞群が選択的に変性・脱落する進行性の難病の総称であり、疾患ごとに固有の異常タンパク質が凝集体を形成して蓄積することが共通の病理学的特徴である。代表的な例としてアルツハイマー病ではアミロイドβやタウ、パーキンソン病(PD)ではαシヌクレイン(αSyn)、筋萎縮性側索硬化症(ALS)ではTDP-43が挙げられる。筆者が研究対象としているPDは神経細胞質内にαSyn凝集体を主要成分とするLewy小体が蓄積することを特徴とする1)。病理学的検討から、Lewy病理は嗅球や延髄から始まり中脳、大脳辺縁系、さらに大脳皮質へと段階的に広がることが示されており(Braak仮説)2)、その進展様式は異常αSynが神経細胞間を伝播し正常αSynを異常構造へと変換しながら脳内に拡大していくというプリオン仮説3)によって説明される。この仮説は培養細胞および動物モデルのいずれにおいても再現されており、異常凝集体が疾患進行の中心的因子であることが確立しつつある。
こうした背景のもと異常凝集体そのものを標的とした抗体医薬や核酸医薬の開発が精力的に進められてきた。アルツハイマー病ではアミロイドβ抗体が臨床応用に至ったものの他の神経変性疾患における開発は依然として困難である4–6)。その主な理由の一つとしてアミロイドβが細胞外に沈着するのに対し、αSynをはじめとする多くの異常凝集体は細胞質内に蓄積するため治療薬の送達が極めて難しいことが挙げられる。
このような課題を踏まえ、筆者らは細胞内に蓄積する異常凝集体を分解する主要な場であるリソソームに着目し、その機能異常と疾患進行との関連を明らかにするとともにリソソーム活性化を基盤とした新たな治療戦略の構築を目指して研究を進めてきた7, 8)。
前述のように異常タンパク質凝集体は神経細胞間を伝播して病変を拡大すると考えられているが、その分子機構の詳細は不明な点が多い。タウやαSynなどの凝集体は細胞外から複数の受容体を介してエンドサイトーシスにより細胞内へ取り込まれることが報告されている9)。しかし、取り込まれた凝集体は分解の場であるリソソームに輸送される。それらは正常タンパクが存在する細胞質とはリソソーム膜によって隔てられているため、「どのようにして細胞外凝集体が細胞質へ到達し内在タンパクの凝集を誘導するのか」は不明であった。近年、αSynを含む神経変性疾患関連の蛋白凝集体が膜傷害性を示すことが報告され10)、我々はリソソーム膜の破綻によって凝集体が細胞質へ漏出し伝播を促進するとの仮説を立てた。
まず、人工的に作製したαSyn凝集体(pre-formed fibril: PFF)をαSyn-EGFPを過剰発現させた細胞に投与し、細胞外凝集体から細胞内タンパクへの凝集伝播の過程を可視化した。PFFは投与後24時間でリソソーム内に高度に集積し(図1-1)、そのうち約5%程度の細胞のリソソームで膜損傷マーカーGalectin-3(Gal3)が陽性、すなわちリソソーム膜の破綻が観察された(図1-2)。タイムラプス撮影ではαSyn-EGFPの凝集がPFFを含むリソソーム近傍から始まる様子が観察され、リソソーム膜破綻部位が凝集伝播の起点であることが示唆された(図1-3、図2)。さらに、このαSyn-EGFP凝集体はヒトLewy小体の主要マーカーであるリン酸化αSyn(pSyn)でも陽性を示し、以後はpSyn免疫染色を指標として内在性αSynの凝集を評価した。pSynはGal3およびLAMP1と共局在し、リソソーム膜を化学的に破綻させる化合物LLOMeにより損傷リソソームを増加させるとPFF投与によるpSyn陽性細胞数が有意に増加した(図1-4)。

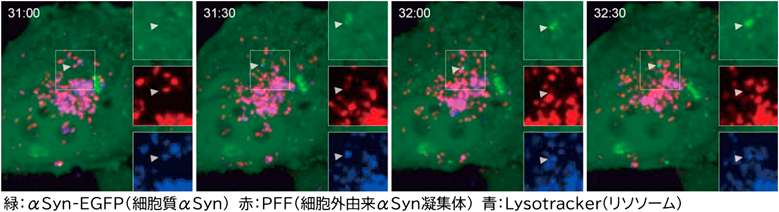
これらの結果から、リソソーム膜が破綻するとその内腔に取り込まれたαSyn凝集体が細胞質へ漏出し内在性αSynの異常凝集を誘導することが明らかとなり、リソソーム膜破綻が異常αSyn伝播の重要な経路であることが示された。
次にPD病態においてリソソーム膜の脆弱性が亢進している可能性に着目した。まず、PDの代表的な遺伝的リスク因子であるGBA1遺伝子について検証した。GBA1はリソソーム酵素グルコセレブロシダーゼをコードしており、その変異はPDの最大の遺伝的危険因子として知られる11)。従来この変異による病態機序として、酵素活性の低下により蓄積するグルコシルセラミドとαSynの相互作用、異常酵素の蓄積による小胞体ストレスなどが報告されていた。我々はGBA1変異によるリソソーム膜脆弱化を検証し、GBA1をノックダウンした細胞では定常状態およびPFF投与後のいずれにおいても野生型細胞に比べてリソソーム損傷細胞(Gal3陽性細胞)が有意に増加した。さらにPFF投与時にはαSyn凝集の伝播(pSyn陽性細胞数)も増加した。これらの結果から、GBA1変異によるリソソーム内部環境の変化12)によりリソソーム膜が脆弱化しαSyn凝集伝播が増強することが示唆された。
一方で、疫学研究からは大気汚染や農薬などの環境因子がPD発症リスクに関与することが知られている。特に大気中の微小粒子状物質(PM2.5)濃度とPD発症率の間に有意な相関が報告され近年注目を集めている13)。我々は金沢大学・瀬戸章文教授の協力を得て大気中から採取されたPM2.5試料を用いた検討を行った。その結果、PM2.5がリソソーム膜傷害能を有すること、特にその構成成分の中でもシリカ粒子が強い膜傷害性を示すことを確認した。さらにPFFとシリカを同時に細胞に投与しすると、広範なリソソーム損傷に伴い細胞質内でのαSyn凝集が有意に促進された。
これらの結果からPD病態において遺伝因子および環境因子が共通してリソソーム膜の脆弱性を高め、異常αSynの伝播を増強することが示された。
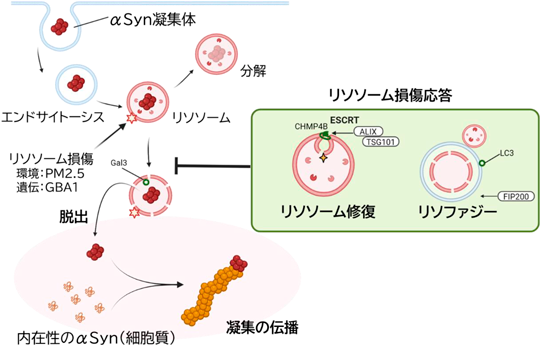
リソソーム膜の脆弱性がパーキンソン病(PD)病態の進展に重要な役割を果たすことが示されたが、損傷を受けたリソソームはリソソーム損傷応答と呼ばれる防御機構により処理されることが知られている14)。この応答は①ESCRT(Endosomal Sorting Complex Required for Transport)複合体による膜損傷部位の修復(リソソーム修復)15, 16)、②損傷リソソームの選択的オートファジーによる隔離・除去(リソファジー)17)、③TFEB核移行シグナルによるリソソームの新生の三つの経路が関与する。これらにより損傷リソソームからの高濃度プロトンや分解酵素等の有害な内容物の細胞質への漏出が防御される。我々はこのリソソーム損傷応答がαSyn凝集体の伝播に対して防御的に働く可能性について検証した。
まずリソファジーに関する検証では、PFFを投与した細胞でLC3のリソソーム上への集積が認められ、電子顕微鏡観察により高電子密度のリソソームが二重膜であるオートファゴソームにより包まれるリソファジーの像を確認した。次にリソファジーが誘導されないFIP200遺伝子ノックアウト細胞(FIP200 KO)を用いて検討を行うと、PFF処理後のFIP200 KO細胞は野生型に比べてリソソーム膜損傷(Gal3)および凝集の伝播(pSyn)が有意に増加した。すなわちリソファジーは損傷リソソームを選択的に隔離・除去することで異常αSynの細胞質漏出および凝集伝播を抑制していることが示された。
続いてESCRT経路による膜修復の寄与を検討した。PFF投与24時間後にはESCRT-IIIサブユニットCHMP4Bのリソソーム上での局在が増加傾向を示し、さらにESCRT関連分子AlixおよびTSG101をダブルノックダウンし修復機構を抑制するとαSyn凝集伝播が有意に増加した。一方でFIP200 KO細胞ではAlix/TSG101ノックダウンによる影響は見られず、リソソーム修復はリソファジーと協調的に機能して有効な防御が成立することが示唆された。
さらにFIP200KO細胞にLLOMeによる高度のリソソーム膜損傷を加えるとαSyn凝集伝播が相乗的に増加した。これらの結果はマウス初代神経細胞を用いた神経細胞間伝播モデルでも再現され、リソソーム損傷応答がリソソーム膜破綻を介するαSyn凝集の細胞間伝播を抑制する主要な防御機構であることが示された18)(図3)。
中枢神経細胞は生涯にわたり分裂・増殖しないため、オートファジー・リソソーム系による細胞内恒常性の維持が極めて重要である。PDを含む神経変性疾患の発症率は加齢とともに著しく上昇し、その背景には老化に伴う有害物質の蓄積によるリソソーム損傷やリソソーム損傷応答の機能低下が関与していると考えられる19)。
我々の研究によりリソソーム損傷と異常タンパク質の蓄積が相互に促進し合う「負のループ」がPDの病態進行の一因であることが示された。また近年、アルツハイマー病など他の神経変性疾患における病的タンパク質であるタウの凝集伝播に対し、リソソーム損傷応答が防御的に機能することが報告された20, 21)。こうした知見からリソソーム損傷応答は異常タンパク凝集体の蓄積を特徴とする神経変性疾患全般に共通する普遍的な防御メカニズムであると考えられる。したがってリソソーム損傷応答の制御およびその活性化は神経変性疾患に対する新たな治療戦略の標的として期待される。現在、筆者らはこのリソソーム損傷応答の詳細な分子機構の解明とその制御による神経保護効果の検証を進めており、これを基盤とした疾患修飾的治療法の創出を目指して研究を推進している。
本稿で紹介した研究成果は大阪大学大学院医学系研究科神経内科学教室で得られたものです。本研究の遂行にあたり、多大なるご指導とご支援を賜りました望月秀樹先生、池中建介先生、大阪大学大学院医学系研究科保健学専攻の吉森保先生、金沢大学微粒子システム研究室の瀬戸章文先生ならびに共同研究者の皆様に心より御礼申し上げます。また、このような栄誉ある賞および執筆の機会を頂戴しました日本神経化学会優秀賞・奨励賞選考委員会および編集委員の先生方に深く感謝申し上げます。今後も研究者として神経化学の発展と神経変性疾患の克服に貢献すべく、これまで以上に研鑽を重ね基礎から臨床へとつながる研究を推進してまいります。
1) Spillantini MG, Schmidt ML, Lee VM, Trojanowski JQ, Jakes R, Goedert M. Alpha-synuclein in Lewy bodies. Nature, 388(6645), 839–840 (1997). doi: 10.1038/42166
2) Braak H, Del Tredici K, Rüb U, de Vos RA, Jansen Steur EN, Braak E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol Aging, 24(2), 197–211 (2003). doi: 10.1016/S0197-4580(02)00065-9
3) Karpowicz RJ Jr., Trojanowski JQ, Lee VM. Transmission of alpha-synuclein seeds in neurodegenerative disease: recent developments. Lab Invest, 99(7), 971–981 (2019). doi: 10.1038/s41374-019-0195-z
4) Lang AE, Siderowf AD, Macklin EA, Poewe W, Brooks DJ, Fernandez HH, Rascol O, Giladi N, Stocchi F, Tanner CM, Postuma RB, Simon DK, Tolosa E, Mollenhauer B, Cedarbaum JM, Fraser K, Xiao J, Evans KC, Graham DL, Sapir I, Inra J, Hutchison RM, Yang M, Fox T, Budd Haeberlein S, Dam T; SPARK Investigators. Trial of cinpanemab in early Parkinson’S disease. N Engl J Med, 387(5), 408–420 (2022). doi: 10.1056/NEJMoa2203395
5) Pagano G, Taylor KI, Anzures-Cabrera J, Marchesi M, Simuni T, Marek K, Postuma RB, Pavese N, Stocchi F, Azulay JP, Mollenhauer B, López-Manzanares L, Russell DS, Boyd JT, Nicholas AP, Luquin MR, Hauser RA, Gasser T, Poewe W, Ricci B, Boulay A, Vogt A, Boess FG, Dukart J, D’Urso G, Finch R, Zanigni S, Monnet A, Pross N, Hahn A, Svoboda H, Britschgi M, Lipsmeier F, Volkova-Volkmar E, Lindemann M, Dziadek S, Holiga Š, Rukina D, Kustermann T, Kerchner GA, Fontoura P, Umbricht D, Doody R, Nikolcheva T, Bonni A; PASADENA Investigators; Prasinezumab Study Group. Trial of prasinezumab in early-stage Parkinson’S disease. N Engl J Med, 387(5), 421–432 (2022). doi: 10.1056/NEJMoa2202867
6) Whone A. Monoclonal antibody therapy in Parkinson’s disease—The End? N Engl J Med, 387(5), 466–467 (2022). doi: 10.1056/NEJMe2207681
7) Oe Y, Kakuda K, Yoshimura SI, Hara N, Hasegawa J, Terawaki S, Kimura Y, Ikenaka K, Suetsugu S, Mochizuki H, Yoshimori T, Nakamura S. PACSIN1 is indispensable for amphisome-lysosome fusion during basal autophagy and subsets of selective autophagy. PLoS Genet, 18(6), e1010264 (2022). doi: 10.1371/journal.pgen.1010264
8) Nakamura S, Shigeyama S, Minami S, Shima T, Akayama S, Matsuda T, Esposito A, Napolitano G, Kuma A, Namba-Hamano T, Nakamura J, Yamamoto K, Sasai M, Tokumura A, Miyamoto M, Oe Y, Fujita T, Terawaki S, Takahashi A, Hamasaki M, Yamamoto M, Okada Y, Komatsu M, Nagai T, Takabatake Y, Xu H, Isaka Y, Ballabio A, Yoshimori T. LC3 lipidation is essential for TFEB activation during the lysosomal damage response to kidney injury. Nat Cell Biol, 22(10), 1252–1263 (2020). doi: 10.1038/s41556-020-00583-9
9) Domingues R, Sant’Anna R, da Fonseca ACC, Robbs BK, Foguel D, Outeiro TF. Extracellular alpha-synuclein: Sensors, receptors, and responses. Neurobiol Dis, 168, 105696–105696 (2022). doi: 10.1016/j.nbd.2022.105696
10) Jiang P, Gan M, Yen SH, McLean PJ, Dickson DW. Impaired endo-lysosomal membrane integrity accelerates the seeding progression of alpha-synuclein aggregates. Sci Rep, 7(1), 7690–7690 (2017). doi: 10.1038/s41598-017-08149-w
11) Nalls MA, Blauwendraat C, Vallerga CL, Heilbron K, Bandres-Ciga S, Chang D, Tan M, Kia DA, Noyce AJ, Xue A, Bras J, Young E, von Coelln R, Simón-Sánchez J, Schulte C, Sharma M, Krohn L, Pihlstrøm L, Siitonen A, Iwaki H, Leonard H, Faghri F, Gibbs JR, Hernandez DG, Scholz SW, Botia JA, Martinez M, Corvol JC, Lesage S, Jankovic J, Shulman LM, Sutherland M, Tienari P, Majamaa K, Toft M, Andreassen OA, Bangale T, Brice A, Yang J, Gan-Or Z, Gasser T, Heutink P, Shulman JM, Wood NW, Hinds DA, Hardy JA, Morris HR, Gratten J, Visscher PM, Graham RR, Singleton AB, Adarmes-Gómez AD, Aguilar M, Aitkulova A, Akhmetzhanov V, Alcalay RN, Alvarez I, Alvarez V, Bandres-Ciga S, Barrero FJ, Bergareche Yarza JA, Bernal-Bernal I, Billingsley K, Blauwendraat C, Blazquez M, Bonilla-Toribio M, Botía JA, Boungiorno MT, Bras J, Brice A, Brockmann K, Bubb V, Buiza-Rueda D, Cámara A, Carrillo F, Carrión-Claro M, Cerdan D, Chelban V, Clarimón J, Clarke C, Compta Y, Cookson MR, Corvol JC, Craig DW, Danjou F, Diez-Fairen M, Dols-Icardo O, Duarte J, Duran R, Escamilla-Sevilla F, Escott-Price V, Ezquerra M, Faghri F, Feliz C, Fernández M, Fernández-Santiago R, Finkbeiner S, Foltynie T, Gan-Or Z, Garcia C, García-Ruiz P, Gasser T, Gibbs JR, Gomez Heredia MJ, Gómez-Garre P, González MM, Gonzalez-Aramburu I, Guelfi S, Guerreiro R, Hardy J, Hassin-Baer S, Hernandez DG, Heutink P, Hoenicka J, Holmans P, Houlden H, Infante J, Iwaki H, Jesús S, Jimenez-Escrig A, Kaishybayeva G, Kaiyrzhanov R, Karimova A, Kia DA, Kinghorn KJ, Koks S, Krohn L, Kulisevsky J, Labrador-Espinosa MA, Leonard HL, Lesage S, Lewis P, Lopez-Sendon JL, Lovering R, Lubbe S, Lungu C, Macias D, Majamaa K, Manzoni C, Marín J, Marinus J, Marti MJ, Martinez M, Martínez Torres I, Martínez-Castrillo JC, Mata M, Mencacci NE, Méndez-del-Barrio C, Middlehurst B, Mínguez A, Mir P, Mok KY, Morris HR, Muñoz E, Nalls MA, Narendra D, Noyce AJ, Ojo OO, Okubadejo NU, Pagola AG, Pastor P, Perez Errazquin F, Periñán-Tocino T, Pihlstrom L, Plun-Favreau H, Quinn J, R’Bibo L, Reed X, Rezola EM, Rizig M, Rizzu P, Robak L, Rodriguez AS, Rouleau GA, Ruiz-Martínez J, Ruz C, Ryten M, Sadykova D, Scholz SW, Schreglmann S, Schulte C, Sharma M, Shashkin C, Shulman JM, Sierra M, Siitonen A, Simón-Sánchez J, Singleton AB, Suarez-Sanmartin E, Taba P, Tabernero C, Tan MX, Tartari JP, Tejera-Parrado C, Toft M, Tolosa E, Trabzuni D, Valldeoriola F, van Hilten JJ, Van Keuren-Jensen K, Vargas-González L, Vela L, Vives F, Williams N, Wood NW, Zharkinbekova N, Zharmukhanov Z, Zholdybayeva E, Zimprich A, Ylikotila P, Shulman LM, von Coelln R, Reich S, Savitt J, Agee M, Alipanahi B, Auton A, Bell RK, Bryc K, Elson SL, Fontanillas P, Furlotte NA, Huber KE, Hicks B, Jewett EM, Jiang Y, Kleinman A, Lin KH, Litterman NK, McCreight JC, McIntyre MH, McManus KF, Mountain JL, Noblin ES, Northover CAM, Pitts SJ, Poznik GD, Sathirapongsasuti JF, Shelton JF, Shringarpure S, Tian C, Tung J, Vacic V, Wang X, Wilson CH, Anderson T, Bentley S, Dalrymple-Alford J, Fowdar J, Gratten J, Halliday G, Henders AK, Hickie I, Kassam I, Kennedy M, Kwok J, Lewis S, Mellick G, Montgomery G, Pearson J, Pitcher T, Sidorenko J, Silburn PA, Vallerga CL, Visscher PM, Wallace L, Wray NR, Xue A, Yang J, Zhang F; 23andMe Research Team; System Genomics of Parkinson’s Disease Consortium; International Parkinson’s Disease Genomics Consortium. Identification of novel risk loci, causal insights, and heritable risk for Parkinson’s disease: A meta-analysis of genome-wide association studies. Lancet Neurol, 18(12), 1091–1102 (2019). doi: 10.1016/S1474-4422(19)30320-5
12) Navarro-Romero A, Fernandez-Gonzalez I, Riera J, Montpeyo M, Albert-Bayo M, Lopez-Royo T, Castillo-Sanchez P, Carnicer-Caceres C, Arranz-Amo JA, Castillo-Ribelles L, Pradas E, Casas J, Vila M, Martinez-Vicente M. Lysosomal lipid alterations caused by glucocerebrosidase deficiency promote lysosomal dysfunction, chaperone-mediated-autophagy deficiency, and alpha-synuclein pathology. NPJ Parkinsons Dis, 8(1), 126 (2022). doi: 10.1038/s41531-022-00397-6
13) Shi L, Wu X, Danesh Yazdi M, Braun D, Abu Awad Y, Wei Y, Liu P, Di Q, Wang Y, Schwartz J, Dominici F, Kioumourtzoglou MA, Zanobetti A. Long-term effects of PM2·5 on neurological disorders in the American Medicare population: a longitudinal cohort study. Lancet Planet Health, 4(12), e557–e565 (2020). doi: 10.1016/S2542-5196(20)30227-8
14) Papadopoulos C, Kravic B, Meyer H. Repair or lysophagy: dealing with damaged Lysosomes. J Mol Biol, 432(1), 231–239 (2020). doi: 10.1016/j.jmb.2019.08.010
15) Radulovic M, Schink KO, Wenzel EM, Nähse V, Bongiovanni A, Lafont F, Stenmark H. ESCRT-mediated lysosome repair precedes lysophagy and promotes cell survival. EMBO J, 37(21), e99753 (2018). doi: 10.15252/embj.201899753
16) Skowyra ML, Schlesinger PH, Naismith TV, Hanson PI. Triggered recruitment of ESCRT machinery promotes endolysosomal repair. Science, 360(6384), eaar5078 (2018). doi: 10.1126/science.aar5078
17) Maejima I, Takahashi A, Omori H, Kimura T, Takabatake Y, Saitoh T, Yamamoto A, Hamasaki M, Noda T, Isaka Y, Yoshimori T. Autophagy sequesters damaged lysosomes to control lysosomal biogenesis and kidney injury. EMBO J, 32(17), 2336–2347 (2013). doi: 10.1038/emboj.2013.171
18) Kakuda K, Ikenaka K, Kuma A, Doi J, Aguirre C, Wang N, Ajiki T, Choong CJ, Kimura Y, Badawy SMM, Shima T, Nakamura S, Baba K, Nagano S, Nagai Y, Yoshimori T, Mochizuki H. Lysophagy protects against propagation of α-synuclein aggregation through ruptured lysosomal vesicles. Proc Natl Acad Sci USA, 121(1), e2312306120 (2024). doi: 10.1073/pnas.2312306120
19) Menzies FM, Fleming A, Caricasole A, Bento CF, Andrews SP, Ashkenazi A, Füllgrabe J, Jackson A, Jimenez Sanchez M, Karabiyik C, Licitra F, Lopez Ramirez A, Pavel M, Puri C, Renna M, Ricketts T, Schlotawa L, Vicinanza M, Won H, Zhu Y, Skidmore J, Rubinsztein DC. Autophagy and Neurodegeneration: Pathogenic Mechanisms and Therapeutic Opportunities. Neuron, 93(5), 1015–1034 (2017). doi: 10.1016/j.neuron.2017.01.022
20) Falcon B, Noad J, McMahon H, Randow F, Goedert M. Galectin-8-mediated selective autophagy protects against seeded tau aggregation. J Biol Chem, 293(7), 2438–2451 (2018). doi: 10.1074/jbc.M117.809293
21) Chen JJ, Nathaniel DL, Raghavan P, Nelson M, Tian R, Tse E, Hong JY, See SK, Mok SA, Hein MY, Southworth DR, Grinberg LT, Gestwicki JE, Leonetti MD, Kampmann M. Compromised function of the ESCRT pathway promotes endolysosomal escape of tau seeds and propagation of tau aggregation. J Biol Chem, 294(50), 18952–18966 (2019). doi: 10.1074/jbc.RA119.009432

This page was created on 2025-11-28T10:40:51.452+09:00
This page was last modified on 2026-01-07T09:20:54.000+09:00
このサイトは(株)国際文献社によって運用されています。